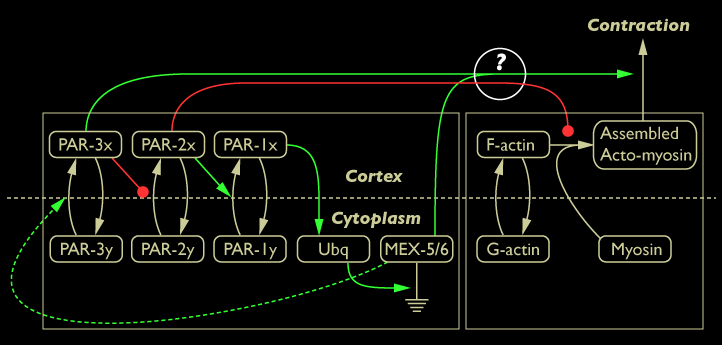
To further characterize the interactions between components of the Par/Mex network and cortical dynamics, we investigated how the cytoplasmic proteins MEX-5/6 influence actomyosin-based contraction of the cortex during the early developmental events leading up to first cleavage in C. elegans. Our interest in MEX-5/6 stems from the observation that in embryos in which MEX-5/6 have been downregulated by RNAi, the clearing of the PAR-3/aPKC-3/PAR-6 complex from the posterior is delayed, not reaching maximal asymmetry until immediatly before first cleavage, suggesting MEX-5/6 promotes rapid clearing of PAR-3/aPKC-3/PAR-6 from the posterior. We wanted to find out whether this effect of MEX-5/6 depletion correlates with a concomitant change in acto-myosin dynamics at the cortex.
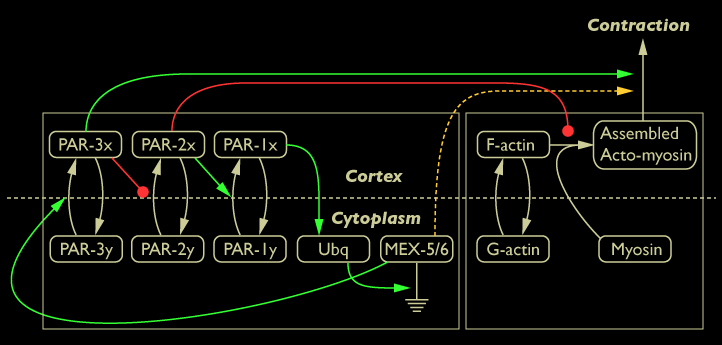
Diagram of interaction between Par/Mex network and cortical actomyosin dynamics (possible link between MEX-5/6 and actomyosin contractility shown in yellow). In addition, although it is not convenient to include it in this diagram, actomyosin contractility leads to asymmetric distribution of Par proteins on the cortex.
We used RNA interference (RNAi) and live confocal imaging to study the affects of MEX-5/6 on actomyosin-based contraction. RNAi is a gene silencing method in which double-stranded RNA is fed to juvenile worms; this targets homologous mRNAs for degradation, effectively knocking down specific genes in the worm's progeny, resulting in aberrant phenotypes. We placed juvenile worms on bacterial feeding plates containing MEX-5/6 double stranded RNA for 24 hours. To examine the effect of downregulating MEX-5/6 on actomyosin-based contraction, we used live confocal microscopy to make time lapse movies of myosin fused with green fluorescent protein. Specifically we were interested in how downregulating MEX-5/6 would affect both global contractility and global cortical flow of the actomyosin meshwork.
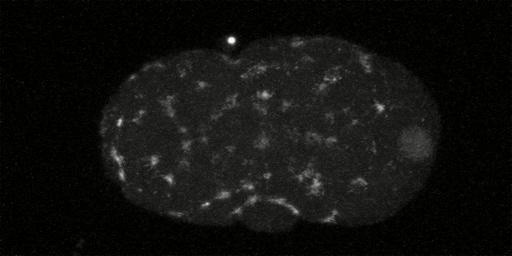
In embryos lacking MEX-5/6, the myosin meshwork in prophase of first cleavage continues to be distributed homgenously along entire cortex at pseudocleavage. At this point in the cell cycle in wild type worms the myosin meshwork has reached maximal asymmetry, having retracted to near 50% embryo length. Anterior is to the left and posterior is to the right. The picture links to a movie [540x480 pixels, 1.7 MB].
Removing MEX-5/6 suppresses global cortical flow of the myosin meshwork seen in wild type embryos [new window, 550x440 pixels, 4.4 MB].; instead we observe a local cortical flow of the myosin meshwork restricted to the posterior cortex. This loss of global cortical flow may explain why we see the myosin meshwork breaking down more posteriorly in embryos lacking MEX-5/6 than in wild-type embryos. To quantify the cortical flow we measured the mean velocity of the myosin foci using kymographs. To analyze myosin foci trajectories we used the program ImageJ to extract subregions from each image of a timelapse sequence and then stack them vertically to make a single kymograph.
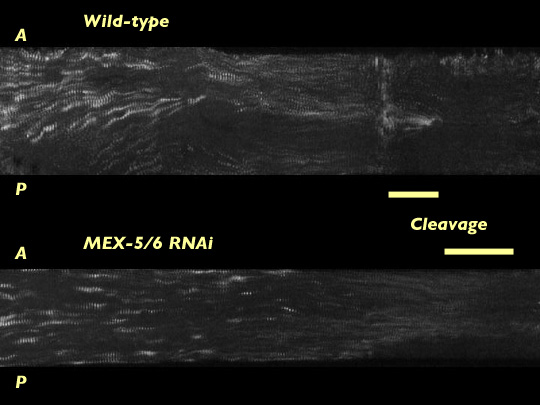
Kymographs constructed from time lapse movies of wild type embryos and embryos lacking MEX-5/6. Time advances left to right.
Mean velocities of myosin foci are greatly reduced in embryos without MEX-5/6. Although there is a local cortical flow at the extreme posterior region, the velocity in this region is less than in wild-type, with velocities of cortical flows in embryos without MEX-5/6 falling off much more rapidly than in wild-type embryos. The reduced velocities of myosin foci indicate MEX-5/6 are an important element in promoting global cortical flow of the myosin meshwork.
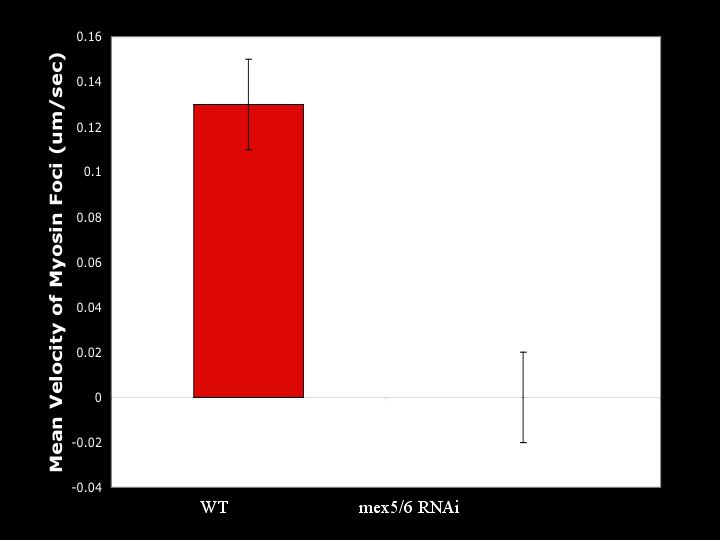
Mean velocity of myosin foci calculations extracted from kymographs.
Removing MEX-5/6 results in the reduction in actomyosin-based contraction of the cortex due to a reduced number of foci. Using ImageJ we quantified the number of foci in wild type embryos and embryos without MEX-5/6.
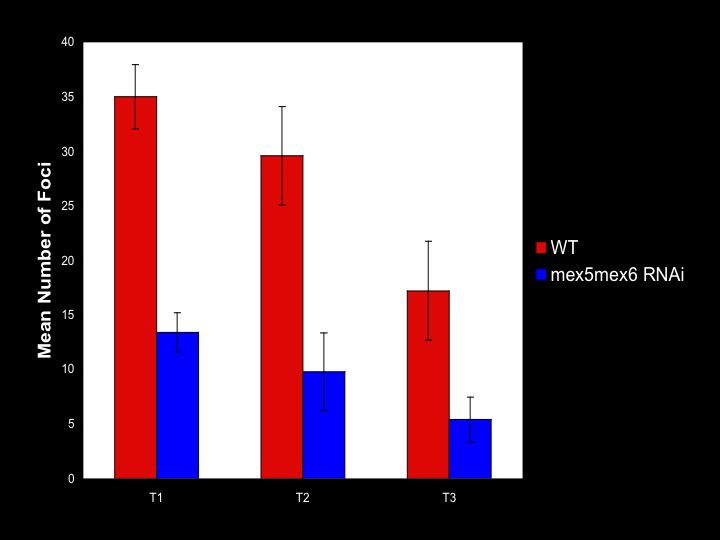
Differences between number of foci in wild type embryos and embryos lacking MEX-5/6 at three different time points in the cell cycle: T1 which corresponds to initial clearing of myosin meshwork from the posterior, T2 corresponding to pseudocleavage, and T3 corresponding to breakdown of the myosin foci.
These results suggest MEX-5/6 may promote the formation of myosin foci. By modulating the number of myosin foci, MEX-5/6 promotes actomyosin-based contraction of the cortex. These two results suggest MEX-5/6 affects global contractile flow and supports the placement of the green line between MEX-5/6 and actomyosin-based contraction of the cortex in the diagram below:
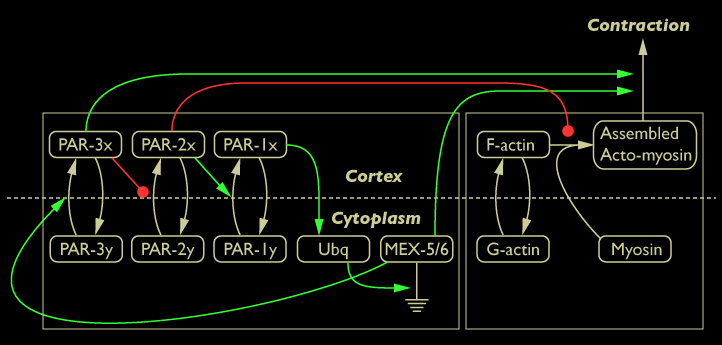
Diagram of interaction between MEX-5/6 proteins and cortical dynamics shown in green
To investigate whether MEX-5/6 is directly affecting actomyosin-based contraction or may be acting indirectly through the actions of PAR-3 (represented by the dotted green line in the network diagram below), we investigated actomyosin-based contraction in par-3 mutants and in par-3 mutants lacking MEX-5/6.
Myosin meshwork in PAR-3 mutant in prophase of first cleavage. The picture links to a movie [550x440 pixels, 3.4 MB]. Movie courtesy of Ed Munro.
As with embryos lacking MEX-5/6, PAR-3 mutants exhibit a local cortical flow of the myosin meshwork in the extreme posterior regions. This lack of global cortical flow of the myosin meshwork is not thought to be due to a decrease in contractility because number of myosin foci appear to be similar to wild type.
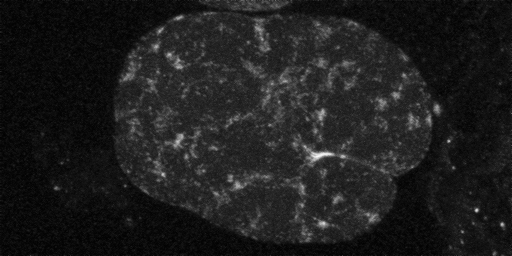
Myosin meshwork in PAR-3 mutant lacking MEX-5/6. View the related movie [opens in a new window, 550x430 pixels, 3.4 MB].
When MEX-5/6 is removed from par-3 mutants, cortical flow is no longer seen. This result suggests both PAR-3 and MEX-5/6 are important components in promoting global contractile flow. These preliminary results suggest PAR-3 and MEX-5/6 may be acting cooperatively to modulate actomyosin-based contraction of the cortex.