Research Assistant
edunn@u.washington.edu
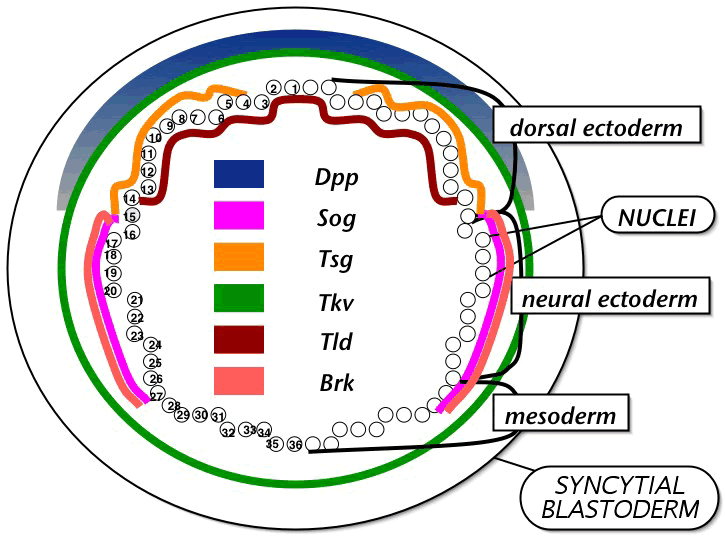
The Drosophila embryo forms a nuclear gradient of maternal dorsal protein after fertilization resulting in a well-documented zygotic gene expression pattern around the circumference of the embryo. My goal is to reveal how gradients of activated proteins interact to create sharp boundaries of gene expression in this system. Nuclear dorsal inhibits transcriptional activation of dpp (decapentapalegic) on the ventral side of the embryo resulting in its uniform expression throughout the dorsal ectoderm only. DPP is a member of the BMP (bone morphogenetic protein) group of TGF signaling molecules. DPP binding to thick veins (TKV) results in phosphorylation of mother against dpp (MAD). DPP is a transcriptional self-activator and it could be necessary for brinker (BRK), produced in the neuroectoderm and an inhibitor of dpp transcription, to keep DPP expression confined to the dorsal ectoderm. DPP protein is observed as a gradient within the dorsal ectoderm. SOG (short gastrulation) is homologous to Xenopus chordin known to antagonize the DPP homolog BMP4. SOG is produced in the neuroectoderm. tld (tolloid) encodes a zinc metalloprotease that cleaves SOG. Twisted gastrulation (TSG) is an accessory protein that enhances the antagonistic activity of SOG, but is required for formation of dorsal-most structures such as the amnioserosa. I am trying to determine (1) how the gradient of DPP is produced (2) how SOG acts as a local inhibitor and long-range activator of DPP and (3) how sog mutant embryos create different patterns of DPP transcriptional targets. I am using the Ingeneue software, developed by Garrett Odell, George von Dassow, Ed Munro and Eli Meir for modeling gene networks, to test models that might explain these phenomena.