
(Brooke Danaher)
Ascidian neurulation proceeds through concurrent invagination and extension of a monolayer neural plate (figure 1). The underlying notochord forms simultaneously through invagination and convergent extension of a monolayer plate into a cylindrical rod (figure 2). The notochord is a likely candidate for effecting neural morphogenesis: it is the substrate for neural tissue throughout neurulation, and its robust convergent extension is a probable source of extensile force in the ascidian tailbud (Munro and Odell, 2002).
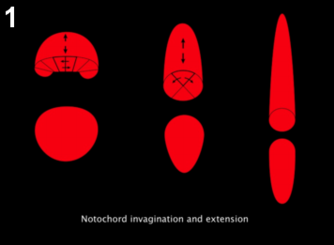
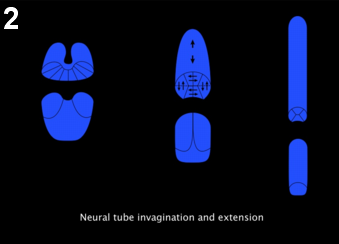
Figure 1: Notochord morphogenesis. At the late gastrula stage (left), notochord cells form a monolayer that intercalate mediolaterally to form a cylindrical rod (middle), that further intercalate to form a single row of cells (right).
Figure 2: Neural tube morphogenesis. At the late gastrula stage, neural plate starts elongating along the antero-posterior axis, while invagination progresses from posterior to anterior (left), forming a cylindrical rod (middle), and subsequently further elongates while the floor and roof cells intercalate giving rise to the typical Ascidian nerve cord with 4 cells in cross section (right).
Here we use laser ablation at the 64-cell stage (figure 3) to examine neural morphogenesis in the absence of a notochord. Using time-lapse microscopy, we show that neural plate organization, invagination, and neural tube closure all continue normally (as shown in movie 1, movie 2, right panel).
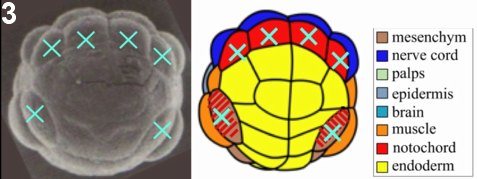
Figure 3: Laser ablation technique. At the 64-cell stage, the 6 precursors of the notochord lineage are ablated. At this stage territories are already determined, so cell fates are not affected by the ablation.
Furthermore, using confocal microscopy of fixed embryos, we show that embryonic overall morphology and tissue organization is surprisingly conserved (figure 4, movie 3, figure 5, movie 4). Neural tube extension is slightly limited, but neural cells retain normal organization and significant elongation (figure 6).
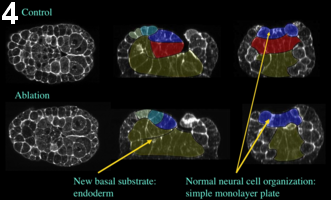
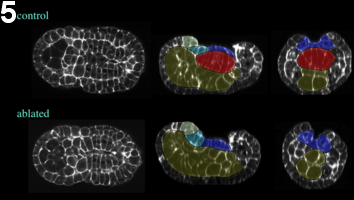
Figure 4: Effect of laser ablation, early neurula stage. Embryos ablated at the 64-cell are fixed at the mid-neurula stage. The general embryonic morphology is preserved. Yellow, endoderm; red, notochord; dark blue, A-line neural cells; light blue and green, posterior and anterior a-line neural cells, respectively. Left, frontal view; middle, sagittal view; right, coronal view.
Figure 5: Effect of laser ablation, early neurula stage. Embryos ablated at the 64-cell are fixed at the late-neurula stage. The general embryonic morphology is preserved. Yellow, endoderm; red, notochord; dark blue, A-line neural cells; light blue and green, posterior and anterior a-line neural cells, respectively. Left, frontal view; middle, sagittal view; right, coronal view.
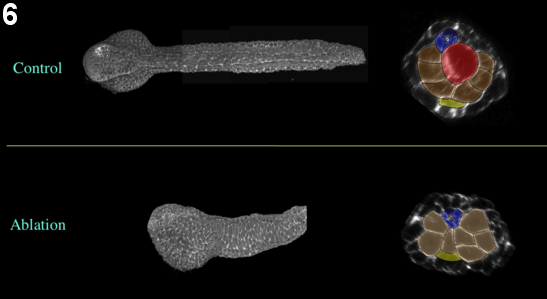
Figure 6: Effect of laser ablation, early neurula stage. Embryos ablated at the 64-cell are fixed at the late tailbud stage. The general embryonic morphology is preserved. Yellow, endoderm; red, notochord; dark blue, A-line neural cells; light blue and green, posterior and anterior a-line neural cells, respectively. Left, frontal view; middle, sagittal view; right, coronal view.
Our results indicate that neural morphogenesis is a robust process, and suggest alternative driving forces for ascidian tailbud elongation.
{kind=link}